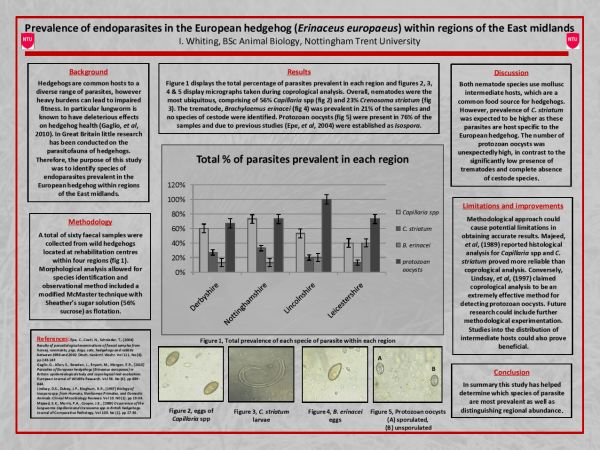
Prevalence of Endoparasites in the European Hedgehog (Erinaceus Europaeus) within Regions of the East Midlands
Isobel Whiting[1], Department of Animal, Rural and Environmental Sciences, Nottingham Trent University
Abstract
Hedgehogs are common hosts to a diverse range of both endo- and ectoparasites (Sykes and Durrant, 2005). This study focused on the prevalence of endoparasites found in the European hedgehog within four regions of the East Midlands. Sixty faecal samples were collected from wild hedgehogs, both males (n=24) and females (n=36) of different ages. The hedgehogs were located at rehabilitation centres in Derbyshire, Nottinghamshire, Lincolnshire and Leicestershire. Faecal samples were analysed using the modified McMaster technique along with Sheather's sugar solution (56% sucrose) as floatation. Parasitic burdens were prevalent in 90% of the samples and morphological analysis allowed for species identification. Nematodes (80%) were the most ubiquitous, comprising of Capillaria spp (56%) and Crenosoma striatum (23%). One species of trematode was identified, Brachylaemus erinacei (21%) and no species of cestode were detected. Protozoan oocysts were present in 76% of the samples and were established as Isospora spp due to previous research (Epe et al., 2004). Lungworm is extremely common among hedgehogs (Boag and Fowler, 1988), as both Capillaria spp and C. striatum use mollusc intermediate hosts, predominantly slugs and snails which are a common food source for hedgehogs (Grewal et al., 2003). However, prevalence of C. striatum was expected to be higher as these parasites are host specific to the European hedgehog (Gaglio et al., 2010). The number of protozoan oocysts was unexpectedly high, in contrast to the significantly low presence of trematodes and complete absence of cestode species. This study helped determine parasitic prevalence of specific species of endoparasites common to the European hedgehog within four regions of the East Midlands.
Keywords: Nematodes, trematodes, protozoan oocysts, endoparasites, European hedgehog, East Midlands.
Introduction
Throughout the United Kingdom (UK) hedgehogs are considered common and widespread; however, studies conducted over the past 20 years show they have been subject to significant decline (Hof and Bright, 2012: 79-88). If this rate of depletion continues research indicates the hedgehog population could become extinct by 2020 (The Mammal Society, 2012). The primary cause of their decline is unclear, although contributing factors include road mortality and anthropogenic disturbances, such as habitat destruction and fragmentation (Dowding et al., 2010a: 13-21). Secondary poisoning due to pesticide applications has also been established as another factor impacting on the species decline (Dowding et al., 2010b: 161-66).
Hedgehogs play an important ecological role due to their position within the food chain. Natural predators include eagles (Accipitridae), stoats (Mustela ermine), polecats (Mustela putorius), foxes (Vulpes vulpes) and more commonly badgers (Meles meles) (The Mammal Society, 2012). In turn, hedgehogs predate a large number of invertebrates and small vertebrates, which has been found to significantly influence localised populations. Therefore, the equilibrium between hedgehog predators and their prey could be detrimentally affected by further decline or potential extinction. Furthermore, hedgehogs serve as a reservoir for a variety of pathogens, therefore loss of this species could consequently cause a significant disparity between parasites and wildlife in the UK (Galglio et al., 2010: 839-44). Additionally a number of these pathogens are zoonotic and therefore carry the risk of transferring disease to humans (Khaldia et al., 2012: 117-22).
Globally there has been limited research conducted on hedgehog parasitofauna and comparatively fewer studies have been conducted throughout the UK (Mizgaiska-Wiktor et al., 2010: 329). Research already undertaken includes the work of Majeed et al., (1989) and Boag and Fowler (1988) who have primarily focussed on the prevalence of lungworm, in particular Capillaria spp and C. striatum. Robinson and Routh (1999) principally directed their research on health implications caused by a number of parasites and pathogens. Lindsay et al., (1997) established high parasite burdens and Sainsbury et al., (1996) recognised the European hedgehog to be an extremely common host for lungworm. Further research into the prevalence of parasites, in particular the pathogenic varieties, could help understand and control wildlife diseases, as well as minimise the risk of potential zoonotic disease transmission (Khaldia et al., 2012: 117-22).
Parasitism
Parasites are highly developmental organisms, evolving to form both direct and indirect lifecycles, whether this be host-specific or to parasitise a variety of hosts (Geyer and Hoffmann, 2012: 221). Parasites are uniquely complex and highly adapted organisms; their numbers exceed that of any free living species, rendering them the most successful and widespread life forms in existence (Krasnov et al., 2004: 1289-97). These diverse organisms are differentiated into two groups, ectoparasites and endoparasites, all of which are completely dependent upon a host organism (Geyer and Hoffmann, 2012: 221). Parasites are generally perceived as damaging, as they exploit a host's resources, resulting in impaired fitness which in turn increases their own fitness. However, a good parasite will not kill its host as it would be destroying the very life source required for survival (Matthews, 1998). Consequently, individuals with impaired fitness are at a higher risk of developing parasitic diseases. This is particularly found to be the case with individuals that are immunosuppressive or who have progressing secondary infections (Boag and Fowler, 1988: 379-82). As previously stated, some parasitic diseases common to the European hedgehog are zoonotic (Khaldia et al., 2011: 117-22). Examples include tick-borne encephalitis (TBE) which is caused by an RNA virus belonging to the genus Flavivirus. Lyme disease is another tick-borne virus from the genus Borrelia (Riley and Chomel, 2005: 2).
Wild species are always at risk of disease, and parasites play an important ecological role in this (Bradley and Altizer, 2007: 97). Eradicating parasites from their hedgehog hosts may not prove viable in the long-term dynamics of a balanced ecosystem as parasites contribute to more than half of life's diversity (Robinson and Routh, 1999: 130).The extent to which parasites damage the health of the European hedgehog is not known. However, it has been reported that they do not appear to be the direct cause of this species' decline (Dowding et al., 2010b: 161-66).
Crenosoma striatum
C. striatum is a parasitic lungworm from the class nematode and is host-specific to the European hedgehog (Boag and Fowler, 1988: 379). C. striatum are heteroxenous parasites, leading an indirect lifecycle (Anderson, 2000). Earthworms (Lumbricus terrestris) and various mollusc species, predominantly land snails (Cepaea hortensis, C. nemoralis and Arianta arbustorum) serve as suitable intermediate hosts (Baruš and Blažek, 1971: 215-26). These molluscs act as paratenic hosts in which transfer of larval stages takes place, allowing the development of the adult stage to occur only within the definitive vertebrate host (Boag and Fowler, 1988: 379-82). This paratenic preference proves to be an effective survival and transmission strategy (Baruš and Blažek, 1971: 215-26). Infection caused by C. striatum typically produces a dry cough, weight loss, nasal discharge, rhinitis, bronchitis followed by possible lesions due to secondary infections. Further symptoms include pulmonary damage leading to pulmonary emphysema or ultimately cardiovascular failure (Boag and Fowler, 1988: 379-82).
Capillaria species
Like C. striatum, Capillaria spp are also found within the class nematode. There are predominantly three species of Capillaria known to infect the European hedgehog (CTD, 2012). C. aerophila is a common parasite of the lung, whereas C. erinacei and C. ovoreticulata parasitise the intestinal tract. Distinguishing between these species can prove difficult due to the strong phenotypic resemblance, therefore they are generically referred to as Capillaria spp (Majeed et al., 1989: 29). In contrast to C. striatum, Capillaria species primarily lead a monoxenous lifecycle, although they have been known to resort to heteroxenous cycles. An infection of C. aerophila occurs when hedgehogs ingest infested organisms such as earthworms which are the primary host (Saupe, 1988: 49-54). Lifecycle is similar to that of C. striatum, in which eggs are coughed up and re-swallowed to then be excreted with faeces (McGarry et al., 1995: 271-72). Symptoms of C. aerophila do not appear to cause as much damage to the respiratory tract as that of C. striatum. Clinical signs usually include irritation of the pulmonary system and coughing. However, secondary bacterial infections commonly arise increasing the risk of pneumonia and further development of parasite burden (Robinson and Routh, 1999: 133). An infection with C. erinacei or C. ovoreticulata is known to affect the intestinal tract. Adult worms' parasitise the intestinal mucosa and as a result infectious lesions can develop (Majeed et al., 1989: 27-36). Symptoms for an infection with C. erinacei or C. ovoreticulata may include chronic enteritis, diarrhoea, weight loss, anaemia and ultimately death (Beck, 2007: 40-44).
Brachylaemus erinacei
B. erinacei is an important digenean trematode from the class trematoda (Carlson, 1990:31-35). The adult form of B. erinacei resides in the hedgehog's intestinal tract; however, heavy burdens will result in migration to the bile ducts (Carlson, 1980: 73-75). The lifecycle of B. erinacei begins when the eggs are shed in the faeces of a definitive host. The eggs then infect a mollusc intermediate host, most commonly terrestrial and water snails, such as various species of Helix (Krehmer, 1967: 524-26). Symptoms of B. erinacei include excessive weight loss along with increased appetite, restlessness and diarrhoea, often containing blood (Robinson and Routh, 1999: 133). More severe cases of infection can lead to haemorrhagic enteritis and inflammation of the bile ducts, along with manifestations of secondary bacterial infections. Such an accumulation of symptoms can lead to anaemia and possibly death (Carlson, 1980: 73-75).
Protozoan oocysts
Protozoan oocysts identified in the European hedgehog have been shown by previous studies (Epe et al., 2004 and Saupe and Poduschka, 1985) to be Isospora species, most commonly I. erinacei, and I. rastegaievae. Certain species from the genus Eimeria have also been reported to infect the European hedgehog (Robinson and Routh, 1999: 131). Isospora and Eimeria can be found in the group coccidia, both of which are intracellular parasites, primarily known for parasitising the intestinal tissues of their host (Robinson and Routh, 1999: 132). Coccidia have complex lifecycles; Isospora in particular express evolutionary monoxenic cycles in which infection occurs upon ingestion of food or water contaminated with sporulated oocysts (Lindsay et al., 1997: 19).
Isospora share a very similar lifecycle to that of Eimeria, the difference being Isospora produce two sporocysts each containing four sporozoites (Duszynski and Upton, 2000: 5). Isospora undergo asexual reproduction by endodyogeny inside the intestinal tract of hedgehogs (Matuschka, 1984: 9-12). This occurs within six days following the initial infection of sporulated oocysts, most of which are in the form of merozoites. Eight days after infection Isospora oocysts are present in the intestinal tract (Robinson and Routh, 1999: 133). Coccidia maintain an effective inter-host transmission strategy aided by a vector, although protozoa generally become encased in a protective cyst. The encysted stage is commonly present in faeces and provides sufficient protection until the next suitable host becomes available (Baker, 1982: 1-24). Hedgehogs infected with Isospora and Eimeria coccidian oocysts will generally appear asymptomatic, although heavy burdens can cause clinical disease which will manifest within the intestinal tract (Epe et al., 2004: 243-47). Clinical signs include weight loss and hematochezia (Robinson and Routh, 1999: 133).
It is apparent that high levels of parasitism occur within the European hedgehog. It has also been established that many of these parasites are vectors for a number of pathogens, many of which will cause disease or impose other deleterious effects within a host (Beck, 2007: 40-44). The small amount of research undertaken in the UK is not current and therefore warrants further investigation within this area.
Aim
The purpose of this study was to determine the prevalence of endoparasites in the European hedgehog within regions of the East Midlands. On a geographical scale there are few studies on the prevalence of parasites in the European hedgehog throughout the UK. Further research establishing parasite prevalence and abundance would aid future investigations and help determine whether there is a significant difference in parasite prevalence dependant on geographic location.
Methodology
Study area
This study took place within four regions of the East Midlands (Figure 1) throughout the months of July, August, September and October 2011. Fifteen faecal samples were collected from each of the four rehabilitation centres (Table 1).
Figure 1: Map of the East Midlands (Source: RTPI, 2011; reproduced by kind permission)
| Rescue centre | Location | Postcode |
| British Hedgehog Preservation | Derbyshire | DE3 9AF |
| Brinsley Animal Rescue | Nottinghamshire | NG16 5AX |
| Weirfields Wildlife Hospital | Lincolnshire | LN6 7PX |
| Helping Hand Hedgehogs | Leicestershire | LE7 3FE |
Table 1: Rehabilitation centres and locations
To ensure faeces remained viable a minimum of five samples were collected at one time. This helped avoid prolonged time periods spent in the lab conducting faecal analysis and also prevented degradation of faeces. Numbers analysed also depended on the availability of faeces from the rehabilitation centres. Hedgehogs in poorer health condition exhibited a loss of appetite, resulting in a reduced production of faeces.
Procedure
Faecal analysis was determined using the McMaster technique along with Sheather's sugar solution. Four grams of faeces were weighed and added to 56ml of floatation fluid. The solution was mixed thoroughly to help break up faecal mass. The resultant was filtered through a muslin membrane into a clean glass beaker, ready to be stirred and withdrawn using a Pasteur pipette. Both chambers of the McMaster slide were filled with the filtration and left for ten minutes to permit floatation. The samples were examined using an Olympus BX51 microscope with an ocular magnification of 4x/10x. Each grid was carefully examined and the number of eggs were recorded and identified. To determine parasitic prevalence, the total number of eggs for each species from both chambers was calculated and multiplied by 50. This method provided an estimate for the total number of eggs per gram (EPG) for each species of parasite. However, to simplify the results the percentage of infected individuals was recorded using presence and absence rather than the number of EPG. During faecal egg analysis, micrographs were taken to help further morphological identification of parasitic egg species as well as support analytical findings. This was achieved using an Olympus BX51-microscope compliant with an Olympus U-CMAD3 adaptable camera.
Results
Of the sixty samples analysed, 90% were found to be burdened with parasites. The most ubiquitous of these parasites were nematodes (80%) in which Capillaria spp were responsible for 56% of infections followed by C. striatum causing 23%. B. erinacei had a total prevalence of 21% and protozoan oocysts comprised 76% of infection. Figure 2 displays the overall results of the study, showing the total prevalence of each parasite within each region.
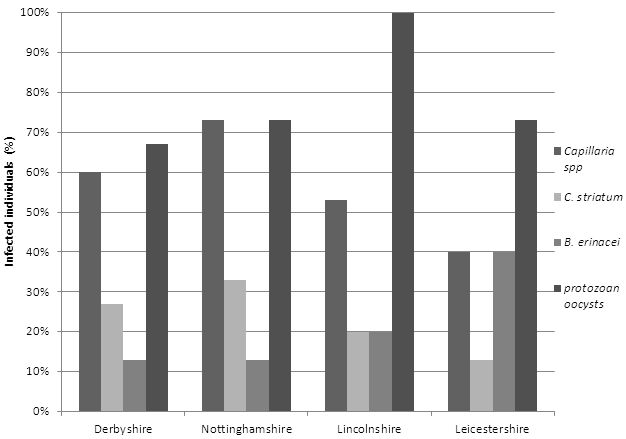
Figure 2: Overall results, showing the percentage of each parasite species within each region.
A comparison between parasite prevalence among male and female hedgehogs was conducted as indicated in figure 3 (n= number).
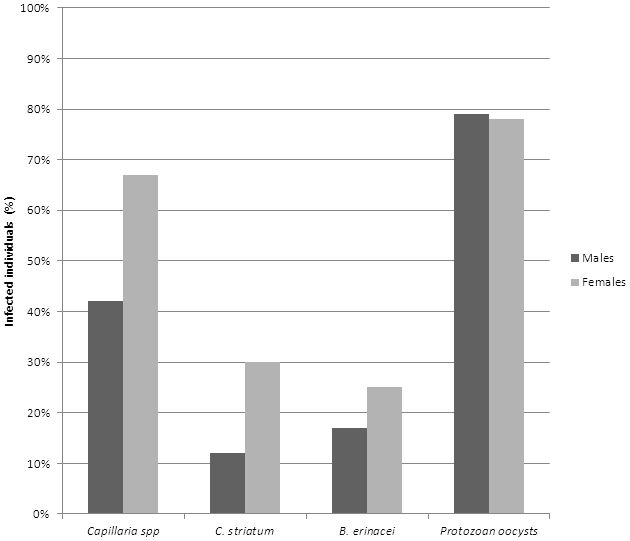
Figure 3: Comparison between male and female hedgehogs. Percentage of parasites prevalent in males (n=24) and females (n=36).
A comparison between parasite prevalence among hedgehogs of differing ages was also conducted as indicated in figure 4.
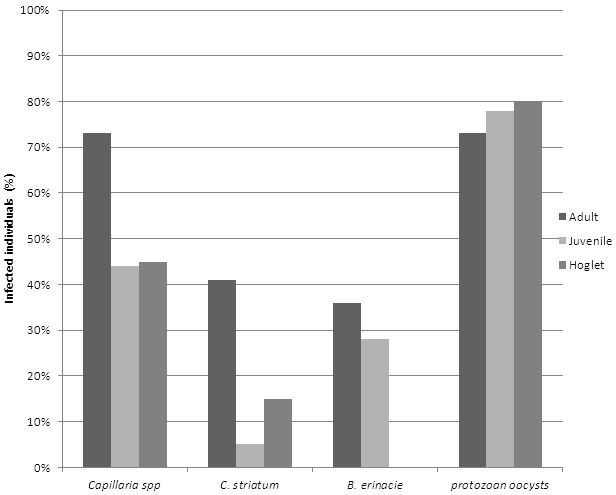
Figure 4: Comparison between hedgehogs of different ages. Percentage of parasites prevalent in the different ages of hedgehogs, inclusive of adults (n=22), juveniles (n=18) and hoglets (n=20).
Morphological Identification
Figures 5 to 8 display micrographs taken during faecal egg analysis. These images help to determine morphological identification of parasite species.
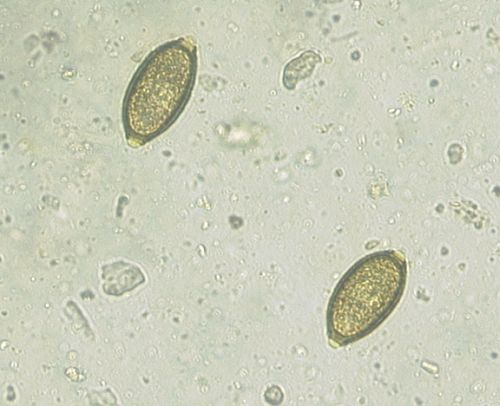
Figure 5: Micrograph displaying the eggs of Capillaria spp.
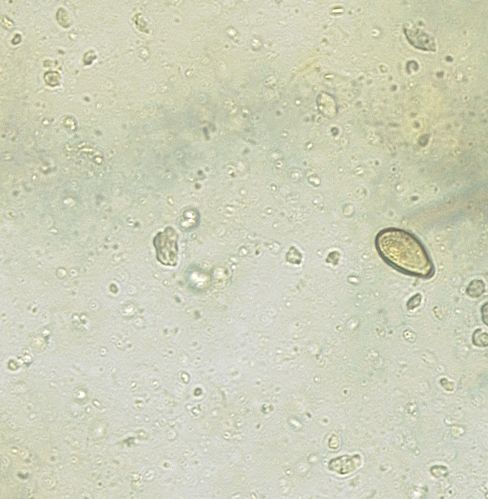
Figure 6: Micrograph displaying an egg of B. erinacei.
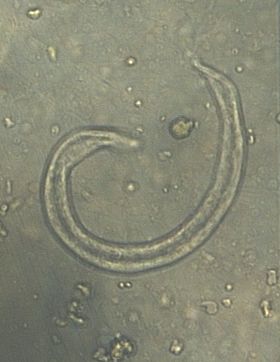
Figure 7: Micrograph displaying the larvae of C. striatum.
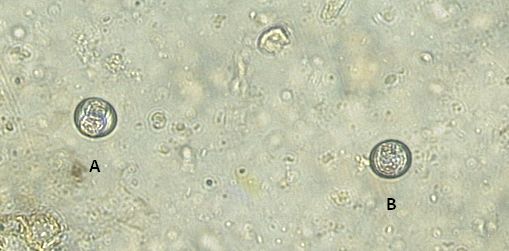
Figure 8: Micrograph displaying protozoan oocysts (A) sporulated (B) unsporulated. This image supports the protozoan oocysts to be Isospora species, as indicated in the sporulated image there are two sporocysts.
Discussion
The aim of this study was to determine the prevalence of endoparasites in the European hedgehog within regions of the East Midlands.
Nematode prevalence
Nottinghamshire was found to have the highest prevalence of Capillaria spp (73%) and C. striatum (33%). The overall prevalence of Capillaria spp was much greater than that of C. striatum. This was unexpected as C. striatum is host-specific to the European hedgehog (Grewal et al., 2003: 146). As previously stated there are three main species of Capillaria known to commonly infect the European hedgehog. Due to the similarity in terms of morphology, it was not possible to identify each species. Therefore, the higher presence of Capillaria to that of C. striatum may have occurred due to the fact that more than one species of Capillaria were detected but for clarification were categorised as Capillaria spp.
Methodology could have influenced the results as reported by Majeed et al., (1989) who found coprological analysis failed to detect up to a third of histologically confirmed Capillaria infestations. The study also proved coprological analysis to be even less reliable at detecting C. striatum. Furthermore, Ciraka et al., (2010) carried out a histological study on parasite prevalence within the southern white breasted hedgehog (E. concolor) in Turkey. Out of the 41 hedgehogs examined C. striatum (72.2%) was observed as having the highest mean intensity of infection. Conversely, Boag and Fowler (1988) undertook a coprological study on parasite prevalence in the UK and found a higher occurrence of Capillaria spp (93%) than C. striatum (77%). Despite being conducted in different countries over a prolonged timescale, these studies could support the report made by Majeed et al. (1989), which leads to the conclusion that histological analysis provides more reliable results than coprological analysis.
Trematode prevalence
The overall prevalence of B. erinacei was significantly low across all four regions compared to the prevalence of nematode species and protozoa. Leicestershire had the highest trematode occurrence in which 40% of the samples had parasites. Robinson and Routh (1999) reported B. erinacei to be more prevalent in southern parts of the UK. Therefore, the low occurrence of B. erinacei established throughout this study could be due to the geographical distribution of the species. It must however be taken into consideration that the report by Robinson and Routh was compiled in 1999. During this study no further research on the geographical distribution of B. erinacei was found to counteract this statement.
Protozoa prevalence
Prevalence of protozoan oocysts was significantly high across all four regions; however, Lincolnshire had 100% prevalence. As previously stated Isospora undergo asexual reproduction within a host's intestinal tract, providing them with an increased ability to reproduce successfully (Matuschka, 1984: 9-12). This factor may help confirm the high prevalence of oocysts detected during this study. The higher occurrence of protozoan may also be due to their effective transmission strategy. As previously stated, protozoa form cysts to ensure protection when vulnerable to the environment (Baker, 1982: 1-24).
Furthermore, Ernst et al. (1986) revealed that the average sporulation time for Isospora oocysts following excretion is approximately 24 hours. This is also dependant on temperature as the incubation period for Isospora requires at least 25°C. A hedgehog's normal body temperature is 35°C, therefore providing a sufficiently warm environment to provoke sporulation. During this study a number of oocysts detected in the faeces were found to have sporulated, the presence of which may account for the high prevalence of protozoa. However, none of the samples acquired during this study should have been older than 24 hours, as each rehabilitation centre confirmed that all enclosures were cleaned on a daily basis. The methodology is an important factor to consider in this instance, as indicated by Lindsay et al., (1997) who claimed Sheather's sugar solution to be an extremely effective method for detecting protozoan oocysts.
Males and females
The sex ratio between males and females had a significant disparity in which females (n=36) exceeded that of males (n=24). This may suggest that the results from the females will provide a more accurate representation of the true occurrence of parasites observed. Despite the females having an overall higher parasitic prevalence, males were found to have a higher occurrence of protozoan oocysts.
According to Isomursu et al. (2006), parasite burdens are more common among male vertebrates. Dimorphism as a result of sexual selection was found to affect an individual's body size, behaviour and immune response. Isomursu et al. (2006) approximated male preference in parasitism occurs due to male sex hormones expressing an immunosuppressive effect. A correlation between body size and parasite burden was also observed. The European hedgehog displays very little sexual dimorphism, although it has been recorded that males are slightly larger than females (Sykes and Durrant, 2005). These findings suggest that the sex of an individual can influence parasite prevalence.
Results from hedgehogs of different ages
Prevalence of parasites among hedgehogs of differing ages found adult hedgehogs to have a higher occurrence of nematode infestation. This could logistically be attributed to a proportionately higher intake of intermediate hosts in comparison to their younger counterparts. In conjunction, older individuals are also more likely to be at risk of infection due to deterioration in immune function (Cichoń et al., 2003: 1206). Furthermore, younger individuals are increasingly susceptible to parasitic infections as their natural defence mechanisms are not fully developed (Morris and English, 1973). The results also showed adult hedgehogs maintain the highest burden of B. erinacei (36%), however the prevalence of this parasite was found to be very low in both adults and juveniles (28%) and were completely absent from the hoglet samples. Unlike the nematode and trematode species the adult hedgehogs had the lowest prevalence of protozoan oocysts (73%). However, this particular parasite was significantly high among the majority of samples.
Robinson and Routh (1999) associated lungworm infection with high morbidity rates in autumn juveniles. This particular study was conducted between July and October. Therefore, some of the individuals could have been autumn juveniles thus contributing towards the overall prevalence of lungworm. However, the results from this study found juveniles to have the lowest occurrence of nematode infections.
Hoglets were found to have the highest prevalence of protozoan oocysts (80%). This could be attributed to transference from mother to offspring as a result of exposure to infected faeces due to small nest size (Matthews, 1998; Bunnell et al., 2011). Hoglets may also be predisposed to parasite burdens due to prenatal transfer. This route of transmission has been recorded in the common canine parasite, Toxocara canis (Matthews, 1998).
Influencing factors
Hedgehogs initiate hibernation between October and November, prior to which feed consumption may have increased in preparation for hibernation. As previously stated, this study was conducted between July and October: consequently, ingestion of infected intermediate hosts may have increased. This will result in a higher prevalence of parasites which subsequently may have influenced the results.
A high proportion of a hedgehog's diet is comprised of invertebrate intermediate hosts, however the exact percentage consumed is unknown (Sykes and Durrant, 2005). A study into the spatial distribution of terrestrial snails was conducted by Chase et al. (1980), in which snail aggregation was observed among individuals of differing ages. Older individuals were found to form clusters more often than younger individuals. The degree of aggregation was also found to be associated with the snails' genetic relationship with preference towards individuals that had hatched from the same cluster. Furthermore, aggregation was found to be less frequent during diurnal hours and more frequent at night. Since hedgehogs are nocturnal it could be suggested that they consume a disproportionately high number of snails due to aggregation, resulting in another factor contributing to parasite prevalence.
A number of studies have been conducted to determine whether there is a correlation between climate change and parasite prevalence. In the UK it was established that the occurrence of Fasciola hepatica, a liver fluke common among sheep, was significantly impacted by rainfall and temperature. As a result the incidence rate of fascioliasis increased considerably (CDC, 2012). Infections with F. hepatica are transmitted via a snail intermediate host (Lymnaea truncatula). Ollerenshaw (1970), established that this transmission is dependent on rainfall and that the size of the snail population increased during wet summers. Foster (1958) reported that the decline in seasonal occurrence of trematode metacercariae present among slugs in the UK was the result of dry and windy conditions, while Svadzhyan (1953) concluded the prevalence of Dicrocoelium lanceatum in molluscs had a greater occurrence in mountain steppe areas. This was due to the frequent occurrence of humid conditions. Conversely, molluscs inhabiting semi desert areas cause prolonged periods of mollusc aestivation, leading to low parasite prevalence. Sub alpine areas may also have lower parasite prevalence due to moderately low temperatures slowing down the development of parasites within the mollusc hosts (Morley and Lewis, 2008: 325–3 35). Similar studies could be conducted to determine a correlation between climate change and parasite prevalence among the European hedgehog.
Future recommendations
Reports indicate the primary reason for hedgehog decline is due to anthropogenic disturbances (Dowding et al., 2010a: 13-21) with no proven indications to suggest any direct correlation between hedgehog decline and parasite burdens (Dowding et al., 2010b: 161-66). As the European hedgehog plays an important role within its ecosystem, future studies could focus on the potential implications of parasite burdens and subsequent hedgehog decline. Such studies could also incorporate a comparison between faecal samples from wild individuals and ones admitted to rehabilitation centres. This information could support hypotheses made regarding higher parasite prevalence among individuals admitted to rehabilitation centres. Conducting malacological surveys throughout the UK would help establish the dispersal and distribution of invertebrates. In addition, such surveys would provide data on the distribution of intermediate hosts.
Hedgehogs pose a potential risk to human health (Khaldia et al., 2011: 117-22). Within rehabilitation centres, carers are continually in direct contact with hedgehogs. Zoonotic diseases are an extreme threat to public health. The most common zoonotic disease spread by hedgehogs in the UK is ringworm (Robinson and Routh, 1999: 132). Keymer et al. (1991) detected ringworm during a hedgehog mortality survey in the UK. Two hedgehogs were also found to have Salmonella typhimurium, while a further thirteen yielded S. enteritidis. Yersinia pseudotuberculosis was also reported in one of the hedgehogs. Each of these pathogens detected are zoonotic, therefore underlining the potential risk to human health. Hedgehogs are becoming increasingly popular as pets and are also the most common wild animal admitted to rehabilitation centres in the UK (The Mammal Society, 2012). Therefore, direct contact with humans is becoming more frequent, thus indicating that further research into hedgehog zoonosis could prove important.
Conclusion
This study investigated the prevalence of endoparasites in the European hedgehog within four regions of the East Midlands. Results found protozoa to have the highest occurrence, comprising of 76% of infection. However, the most ubiquitous parasite was found to be nematodes (80%) inclusive of Capillaria spp (56%) and C. striatum (23%). The trematode, B. erinacei maintained the lowest occurrence of 21%. It was also established that gender and age have an important role in the prevalence of parasites (Isomursu et al., 2006; Cichoń et al., 2003). Recent research into hedgehog parasitofauna in the UK is limited although findings from this study support similar findings from previous studies, despite there being a disparity in time period.
Previous research into the impact of parasite prevalence and climatic conditions have shown there to be a correlation among a number of species (Polley et al., 2010; Morley and Lewis, 2008; Jenkins et al., 2006; Haukisalmi and Henttonen, 1990; Ollerenshaw, 1970; Foster, 1958; Svadzhyan, 1953). This information could be applied to parasites common to the European hedgehog. The importance of intermediate hosts has also been highlighted during this study, as the European hedgehog's diet comprises primarily of invertebrates, in particular molluscs (Morris, 2006). The abundance and distribution of these species is extremely important when considering parasite transmission to the definitive host (Jenkins et al., 2006: 387-401). During this study insufficient data was found on both the population levels and dispersal of invertebrates throughout the UK. However, the main limitation of this study was found to be the experimental method used for faecal egg analysis. Despite previous parasitological studies reporting coprological analysis to be an effective method (Lindsay et al., 1997; Boag and Fowler, 1988), other studies claimed histological analysis to provide more reliable results, particularly when detecting C. striatum. In the opinion of Majeed et al. (1989), faecal analysis is inaccurate and can fail to detect certain species of parasite. The potential risk of hedgehog zoonosis in relation to public health warrants further investigation. Findings by Keymer et al. (1991) support the need for such studies as various zoonotic pathogens were found within the European hedgehog in the UK.
Overall, this study represents a comparative investigation into the prevalence of certain species of endoparasites common to the European hedgehog within regions of the East Midlands. Findings from the study confirm hedgehogs are commonly parasitised and carry significantly high burdens of parasites, in particular nematodes and protozoa.
Acknowledgements
I would like to express my appreciation to Dr Jacqueline Boyd for her continued guidance and supervision throughout this research. I am also extremely grateful to the rehabilitation centres for providing me with the samples required for this study and to the laboratory staff at NTU for their assistance during faecal analysis. Additionally, I would like to thank Dawn Mackie for proofreading my thesis and providing me with constructive feedback and finally I would like to thank Tom Wah Brannigan for providing his appraisals and criticisms from a non-scientific perspective.
List of Figures
Figure 1: Map of the East Midlands (Source: RTPI, 2011; reproduced by kind permission)
Figure 2: Overall results
Figure 3: Comparison between male and female hedgehogs
Figure 4: Comparison between hedgehogs of different ages
Figure 5: Micrograph displaying the eggs of Capillaria spp
Figure 6: Micrograph displaying the eggs of B. erinacei
Figure 7: Micrograph displaying the larvae of C. striatum
Figure 8: Micrograph displaying protozoan oocysts (A) sporulated (B) unsporulated.
List of Tables
Table 1: Rehabilitation centres and locations
Notes
[1] Isobel Whiting graduated from Nottingham Trent University with a 2:1 honours degree in Animal Biology and is looking to develop a career in research.
References
Anderson, R. C. (2000), Nematode parasites of vertebrates: Their development and transmission, UK: CABI Publishing
Baruš, V. and K. Blažek (1971), 'The life cycle and the pathogenicity of the nematode Crenosoma striatum', Folia Parasitologica, 18, 215-26
Beck, W. (2007), 'Endoparasiten beim Igel. Wiener Klinische Wochenschrift'< /font>, The Middle European Journal of Medicine, 119, 40-44
Baker, J. R. (1982), Parasitic Protozoa in British Wild Animals, Cambridge: Institute of Terrestrial Ecology
Boag, B. and P. A. Fowler (1988), 'The prevalence of helminth parasites from the hedgehog Erinaceus europaeus in Great Britain', Journal of Zoology, 215, 379-82
Bradley, C. A. and S. Altizer (2007), 'Urbanization and the ecology of wildlife diseases', Trends in Ecology and Evolution, 22 (2), 95-102
Bunnell, T., K. Hanisch, J.D. Hardegeand T. Breithaupt(2011), 'The Faecal Odour of Sick Hedgehogs (Erinaceus europaeus) Mediates Olfactory Attraction of the Tick Ixodes hexagonus', Journal of Chemical Ecology, 37(4), 340-47
Carlson, A. (1980) 'Diagnose und Therapie der Parasitosen der Igel', Der Praktische Tierarzt, 62, 73-75
Carlson, A. (1990), 'Der Igel in der tierärztlichen Praxis', Der Praktische Tierarzt, 70, 31-35
Centre for Disease Control (CDC), (2012), 'Parasites of the Intestinal Tract', available at http://dpd.cdc.gov/dpdx/html/Para_Health.htm, accessed 5January 2012
Chase, R., R. P. Croll and L. L. Zeichner (1980), 'Aggregation in Snails, Achatina fulica', Behavioural and Neural Biology, 30, 218-30
Cichoń, M., J. Sendecka and L. Gustafsson (2003), 'Age-related decline in humeral immune function in Collared Flycatchers', Journal of Evolutionary Biology, 16 (6), 1205-10
Ciraka, V. Y., B. Senlika, A. Aydogdub, M. Selvera and V. Akyola (2010), 'Helminth parasites found in hedgehogs (Erinaceus concolor) from Turkey', Preventive Veterinary Medicine, 97, 64-66
Comparative Toxicogenomics Database (CTD), (2012), 'Ancestor Path', available at http://ctdbase.org, accessed 2 January 2012
Dowding, C. V., S. Harris, S. Poulton and P. J. Baker (2010a), 'Nocturnal ranging behaviour of urban hedgehogs, Erinaceus europaeus, in relation to risk and reward', Animal Behaviour, 80, 13-21
Dowding, C. V., R. F. Shore, A. Worgan, P. J. Baker and S. Harris (2010b), 'Accumulation of anticoagulant rodenticides in a non-target insectivore, the European hedgehog (Erinaceus europaeus)', Environmental Pollution, 158 (1), 161–66
DuszynskI, D. W. and S. J. Upton (2000), 'Coccidia (Acomplexa: Eimeriidae) of the Mammalian Order Insectivora', The Museum of Southwestern Biology, 4, 1-67
Ernst, J. V., D. S. Lindsay, J. A. Jarvinen, K. S. Todd and D. P. Bane (1986), 'The sporulation time of Isospora suis oocysts from different sources', Veterinary Parasitology, 22 (1-2), 1-8
Epe, C., N. Coati and T. Schneider (2004), 'Results of parasitological examinations of faecal samples from horses, ruminants, pigs, dogs, cats, hedgehogs and rabbits between 1998 and 2002', Deutsche tierärztliche Wochenschrift, 111 (6), 243-47
Foster, R. (1958), 'Infestation of the slugs Milax sowerbii Ferussac and Agriolimax reticulatus Muller by trematode metacercariae (Brachylaemidae)', Parasitology, 48, 303-11
Gaglio, G., S. Allen, L. Bowden, M. Bryant and E. R. Morgan (2010), 'Parasites of European hedgehogs (Erinaceus europaeus) in Britain: epidemiological study and coprological test evaluation', European Journal of Wildlife Research, 56 (6), 839-44
Geyer, K. K. and K. F. Hoffmann (2012), 'Epigenetics: A key regulator of platyhelminth developmental biology', International Journal of Parasitology, 42 (3), 221-24
Grewal, P. S., S .K. Grewal, L. Tan and B. J. Adams (2003), 'Parasitism of Molluscs by Nematodes: Types of Associations and Evolutionary Trends' Journal of Nematology, 35 (2), 146-56
Haukisalmi, V. and H. Henttonen(1990), 'The impact of climatic factors and host density on the long-term population dynamics of vole helminths', Oecologia, 83 (3), 309-15
Hof, A. R. and P. W. Bright(2012), 'Factors affecting hedgehog presence on farmland as assessed by a questionnaire survey', Acta Theriologica, 57 (1), 79-88
Isomursu, M., O. Rätti, P. Helle and T. Hollmén (2006), 'Sex and age influence intestinal parasite burden in three boreal grouse species', Journal of Avian Biology, 37 (5), 516-22
Jenkins, E. J., A .M. Veitch, S. J. Kutz, E. P. Hoberg and L. Polley (2006), 'Climate change and the epidemiology of protostrongylid nematodes in northern ecosystems: Parelaphostrongylus odocoilei and Protostrongylus stilesi in Dall's sheep (Ovis d. dalli)', Parasitology, 132 (3), 387-401
Khaldia, M., C. Socolovschib, M. Benyettoua, G. Barecha, M. Bichec, T. Kernifb, D. Raoultb and P. Parolab (2012), 'Rickettsiae in arthropods collected from the North African Hedgehog (Atelerix algirus) and the desert hedgehog (Paraechinus aethiopicus) in Algeria', Comparative Immunology, Microbiology and Infectious Diseases, 35 (2), 117-22
Krasnov, B. R., G. I. Shenbrot, I. S. Khokhlova and R. Poulin (2004), 'Relationships between parasite abundance and the taxonomic distance among a parasite's host species: an example with fleas parasitic on small mammals', Journal of parasitology, 34 (11), 1289-97
Krehmer, E. (1967), 'Starker Befall mit Brachylaemus erinacei (Trematoda: Brachylaemidae) als Todesursache eines Igels', Tierärztliche Umschau, 22, 524-26
Lindsay, D. S., J. P. Dubey and B. R. Blagburn (1997) 'Biology of Isospora spp. from Humans, Nonhuman Primates, and Domestic Animals', Clinical Microbiology Reviews, 10 (1), 19-34
Majeed, S. K., P. A. Morris and J. E. Cooper (1989), 'Occurrence of the lungworms Capillaria and Crenosoma spp in British hedgehogs', Journal of Comparative Pathology, 100 (1), 27-36
Mammal Society, (2012), 'Hedgehog Survey Pilot 2009', available at http://www.mammal.org.uk/index.php?option=com_content&view=article&id=310&Itemid=344, accessed 1 February 2012
Matthews, B. E. (1998), An Introduction to parasitology, Cambridge: Cambridge University Press
Matuschka, F. R. (1984), 'Endogeny in Isospora rastegaievae from the Eurasian hedgehog (Erinaceus europaeus L.)', Parasitology, 88, 9-12
McGarry, J. W., M. Martion, M. T. Cheeseman and C. E. Payne-Jophnson (1995), 'Crenosoma vulpis, the fox lungworm, in dogs', Veterinary Record, 137, 271-72
Mizgaiska-Wiktor, H., W. Jarosz, B. Piiłacińska and S. Dziemian (2010), 'Helminths of the hedgehog, Erinaceus europaeus and E. roumanicus from Poznań region, Poland—coprological study', Wiadomo ści parazytologiczne, 56 (4), 329-32
Morris, P. (2006), The New Hedgehog Book, Suffolk: Whittet Books
Morris, P. and M. P. English (1973), 'Transmission and course of Trichophytonerinacei in British hedgehogs', Medical Mycology, 11 (1), 42-47
Morley, N. J. and J. W. Lewis (2008), 'The influence of climatic conditions on long-term changes in the helminth fauna of terrestrial molluscs and the implications for parasite transmission in southern England', Journal of Helminthology, 82, 325–3 5
Ollerenshaw, C. B. (1970), 'Some aspects of the epidemiology of Fascioliasis in Britain', Proceedings of the Malacological Societ, 39, 238–43
Polley, L., E. Hoberg and S. Kutz (2010), 'Climate change, parasites and shifting boundaries', ACTA Veterinaria Scandinavica, 52 (1), 51-55
Riley, P. Y. and B. B. Chomel (2005), 'Hedgehog Zoonosis', Emerging Infectious Diseases, 11 (1), 1-5
Robinson, I. and A. Routh (1999), 'Veterinary care of hedgehogs' Journal of the British Veterinary Association. In Practice, 21, 128-37
RTPI (Royal Town Planning Institute), (2011), 'Maps of East Midland', available at http://www.rtpi.org.uk/knowledge/networks/young-planners-network/young-planners-in-your-area/east-midlands-young-planners/, accessed 1 September 2011
Sainsbury, A. W., A. A. Cunningham, P. A. Morris, J. K. Kirkwood and S. K. Macgregor (1996), 'Health and welfare of rehabilitated juvenile hedgehogs (Erinaceus europaeus) before and after release into the wild', Veterinary Records, 138, 61-63
Saupe, E. and W. Poduschka (1985), 'Hedgehog', Diseases of Companion Animals, 75-96
Saupe, E. (1988), 'Die Parasitosen des Igels und ihre Behandlung', Der Praktische Tierarzt, 12, 49-54
Svadzhyan, P. K. (1953), Dynamics of infestation of terrestrial molluscs of Armenian S.S.R. by parthenogenetic stages of Dicrocoelium lanceatum and factors influencing the emergence of multiple cysts, Petrov
Sykes, L. and J. Durrant (2005), The Natural Hedgehog, London: Gaia Books
To cite this paper please use the following details: Whiting, I. (2012), 'Prevalence of Endoparasites in the European Hedgehog (Erinaceus Europaeus) within Regions of the East Midlands', Reinvention: a Journal of Undergraduate Research, British Conference of Undergraduate Research 2012 Special Issue, www.warwick.ac.uk/go/reinventionjournal/issues/bcur2012specialissue/whiting. Date accessed [insert date]. If you cite this article or use it in any teaching or other related activities please let us know by e-mailing us at Reinventionjournal@warwick.ac.uk.
Poster from BCUR
Click on image to download full PDF version