
Evolution
Evolution is one of the central tenets of biology. In essense it answers the question "why?": the reason why do organisms behave, look or interact in the ways that they do is the consequence of evolution. Work in this group is interested in understanding evolution of infectious diseases. In particular we wish to address three main questions,
1) Can the observed parameters associated with diseases be explained by evolutionary models?
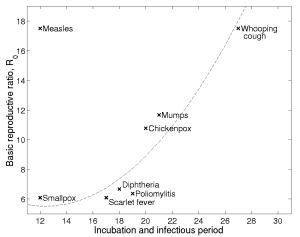 |
This diagram shows two important characteristics associated with several human airborne pathogens (the x-axis shows the average amount of time an individual is infected while the y-axis shows the basic reproductive ratio -- a measure of the diseases absolute infectivity). It is clear that many pathogens lie close to the dashed curve, such that increasing length of time infected is associated with an increasing potential to generate more secondary cases -- although the relationship is clearly non-linear. Measles however is an outlier. Why are its parameter values so different? Are there evolutionary explanations for contraint of paramaters by the dashed curve? Are there reasons why measles has escaped this constraint? |
2) What are the implications of recent changes in human social mixing for the evolution of infection diseases?
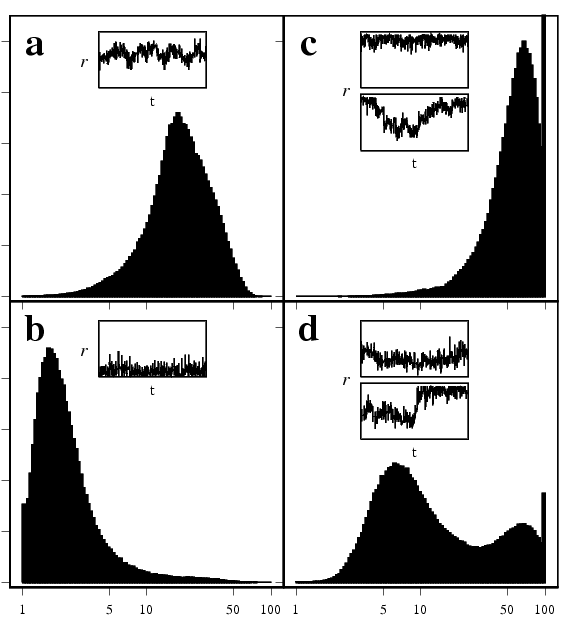 It is reasonable well established that over the past century there has been a dramatic change in the social interactions of many individuals. In the 1900's most people lived and worked within a small village and rarely travelled long distances. Today, long-distance commuting is common-place, busness travellers fly all over the world for meeting and long-haul holidays are becoming the norm. Clearly these changes have strongly influenced the network of contacts through which infectious diseases spread. We can examine the impacts of such network structure on the long-term evolutionationary dynamics of diseases (see the figure on the right). Two different networks (local and global) and two different evolutionary constraints are considered; clearly both are vitally important to understand disease evolution.
It is reasonable well established that over the past century there has been a dramatic change in the social interactions of many individuals. In the 1900's most people lived and worked within a small village and rarely travelled long distances. Today, long-distance commuting is common-place, busness travellers fly all over the world for meeting and long-haul holidays are becoming the norm. Clearly these changes have strongly influenced the network of contacts through which infectious diseases spread. We can examine the impacts of such network structure on the long-term evolutionationary dynamics of diseases (see the figure on the right). Two different networks (local and global) and two different evolutionary constraints are considered; clearly both are vitally important to understand disease evolution.
3) What constraints on evolution are due to the interaction of the pathogen with the immune system?
In simple models of disease evolution a relationship is often imposed between the transmission rate (β) and the duration of infection (1/γ). If formulated correctly this trade-off can prevent run-away evolution to ever higher parameter values and can lead to intermediate evolutionarily stable strategies. Are such relationships realistic and can they be predicted from models of within-host dynamics?
Publications
JM Read, MJ Keeling (2007) Stochasticity generates an evolutionary instability for infectious disease. Ecology Letters (in press).
JM Read, MJ Keeling (2006) Disease evolution across a range of spatio-temporal scales. Theo. Pop. Biol. 70, 201-213
KTD Eames, MJ Keeling (2006) Coexistence and specialization of pathogen strains on contact networks. Am. Nat. 168, 230-241
MJ Keeling, F Jiggins, JM Read (2003). The invasion and coexistence of competing Wolbachia strains. Heredity 91, 382-388.
JM Read, MJ Keeling (2003). Disease evolution on networks: the role of contact structure. Proc. Roy. Soc. Lond. B 270, 699-708.
Keeling (2000) Evolutionary Trade-Offs at Two Time Scales: Competition v Persistence Proc. Roy. Soc. Lond. B 267 385-391
Rand, Keeling and Wilson (1995) Invasion, Stability and Evolution to Criticality in Spatially Extended Artificial Host-Pathogen Ecologies Proc. R. Soc. Lond. B 259 55-63
Keeling and Rand (1995) A Spatial Mechanism for the Evolution and Maintenance of Sexual Reproduction Oikos 74 414-424
Funded by:
NIH

NSERC

Leverhulme Trust

People involved:
Matt Keeling
Jane Heffernan
Jon Read
Mike Boots (Sheffield)
Steve Webb (Strathclyde)