
Phylogeography
Allaby Research Group
Phylogeography uses the distributions of alleles from extant plant accessions to establish past movement patterns. We use the term 'modogenetics' to discriminate this type of genetic data from archaeogenetics, modo- being the Latin root for 'just now' (as opposed to neo- which means 'new', but usually has connotations of resurgence rather than extant). Our activities in ecological genomics would also be classed as modogenetics.
Human mediated movement and local adaptation
It has been a general observation over the past couple of decades that the phylogeographic distributions of many plant species, particularly those that are considered more 'primitive' (old landraces of globally important plants or plants which are not of primary global importance), appear to reflect ancient human population movements. An example is described below with maize, similar findings have been made with emmer wheat (Brown et al 2006). This is fascinating on two counts. Firstly, it provides an insight into early human activities - the current example we are engaged in concerning the taro crop in Africa is outlined below. Secondly, it suggests that some of these plant populations have been in situ for very long periods of time (thousands of years in some cases) after being moved out of their centre of origin. Such populations should be locally adapted to their immediate environment. This is of global significance given the increased food production that will be required in the 21st century. Ancient civilizations may have done a lot of our science for us through breeding programmes on a scale unimaginable in modern science. If we can discover what makes, or indeed made, crops suited to local conditions such as ones under high drought stress, or drought and cold that might be experienced under high altitude conditions, then we may be able to introduce such qualities into our modern crop stocks. Consequently a major thrust of our research is focussed on discovering local adaptation on a genome wide level. This through archaeogenetic approaches (see barley), and through natural wild populations (see ecological genomics).
Taro (Colocasia esculenta) in Africa, and the SEALINKS project
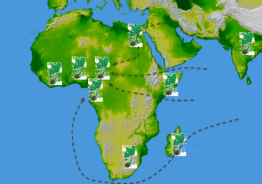

Taro is one three crops of south east Asian origin (the others are banana and yam, see Allaby 2007) that are found in Africa. It is currently thought that these crops probably moved together, transported by early seafarers several thousand years ago. Taro seems to have been domesticated in several centres, including south east Asia, Papua New Guinea, and around the Assam region. The aim of this project is to determine early seafaring activity by tracing the mode of entry into Africa of taro. It could have entered on the east cast, brought by Polynesian peoples, it could have been brought more directly (and possibly much earlier) more directly from South Asia, over land or following the coast, or it might have been brought around the African continent to the Cameroon area, where it has the highest cultivar diversity today. All hypotheses are not without problems. By examining chloroplast and microsatellite data we aim to see which of these hypotheses has more credence, which in turn will tell us not only about taro, but about the people who brought taro to Africa.
Phylogeography of South American maize
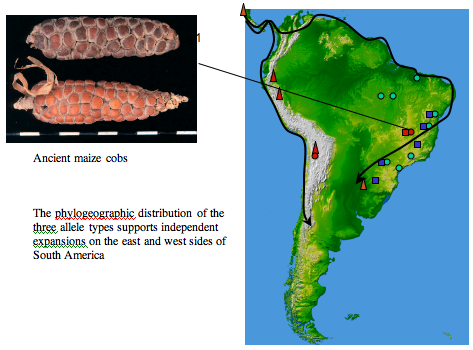
In this case primitive maize landraces showed a distinct patterning of allelic distribution across the South American continent which appeared to reflect to routes of entry from Mesoamerica. Interestingly, this allelic pattern is also reflected in archaeological samples of maize collected from Andean regions and the lowland tropics of Brazil (Freitas et al 2003). This work is ongoing in the Manchester ancient DNA group.
Phylogeography publications
Allaby, R.G. (2007) Origins of Plant Exploitation in the Pacific: A review. In Population Genetics, Linguistics, and Culture History in the Southwest Pacific: A Synthesis. Ed. Friedlaender, J.F., OUP.
Brown T.A., Lindsay S., and Allaby R.G. (2006) Using modern landraces of wheat to study the origins of European Agriculture. In Darwin's Harvest. Pp. 197-212. Eds. Motley, T.J., Zeregra, N., Cross, H. Columbia University Press, New York.
Allaby, R.G., Peterson, G., Merriwether, D.A., Fu, Y.-Bi (2005) Evidence of the domestication history of flax (Linum usitatissimum L.) from genetic diversity of the sad2 locus. Theor. Appl. Genet. 112: 58-65.
Freitas F.O., Bendel G., Allaby R.G., and Terence A. Brown (2003) DNA from Primitive Maize Landraces and Archaeological Remains: Implications for the Domestication of Maize and its Expansion into South America. Journal of Archaeological Science 30:901-908.
Allaby R.G. and Brown T.A. (2001) Network analysis provides insights into the evolution of 5S rDNA arrays in Triticum and Aegilops. Genetics 157:1331-1341.
Allaby R.G. and Brown T.A. (2000) Identification of a 5S rDNA spacer type specific to Triticum urartu and wheats containing the T. urartu genome. Genome 43:250-254.
Allaby R.G., Banerjee M., Brown T.A. (1999). Evolution of the high molecular weight glutenin loci of the A,B,D and G genomes of wheat. Genome 42:296-307.
Jones M.K., Brown T.A., Allaby R.G. (1997) Wheat domestication. Science 279:302-303.
Jones M.K., Brown T.A., Allaby R.G. (1996). Tracking early crops and early farmers: the potential of biomolecular archaeology. In The Origins and Spread of Agriculture and Pastoralism in Eurasia. (ed. D.R. Harris) UCL press, London.